INTRODUCTION
The widespread application of electric (often called artificial) light in greenhouses, growing rooms, and plant growth chambers would presuppose that the role of phytochrome would be considered in the selection and use of such lighting systems. Unfortunately this is not usually the case. Part of the problem is that many students, and indeed an unfortunate number of senior scientists, seem to regard phytochrome as a laboratory phenomenon without much application in the real world. They simply have not grasped the concept that phytochrome is functioning through all stages of plant development, wherever plants are grown. It is certainly true, as Meijer (1971) stated, that one cannot compare experimental results obtained under very strict laboratory conditions with plant irradiation in glasshouses and in growth rooms. For example, the action spectrum for flowering of the long-day plant, Hyoscyamus niger, (Parker et al., 1950) clearly shows that red radiation is the most efficient portion of the spectrum for promoting flower initiation, but in practical photoperiod control red or fluorescent lamps do not promote flowering nearly as well as the mixture of red and far-red in incandescent lamps. Nevertheless, much evidence exists that documents phytochrome control of plant growth and development in controlled environments and under natural conditions.
When Karl Norris developed the first practical portable spectroradiometer about 1962 some of the first measurements were to determine the red/far-red ratios under tree canopies (Downs and Hellmers, 1975). These measurements showed clearly the predominance of far-red in the understory and suggested that far-red was contributing to the elongation exhibited by many species growing in the shade, and possibly was a factor in the induction of light requirements in seeds. Subsequently we used Catalpa leaves as far-red filters to make light-insensitive lettuce seed light requiring. Much more detailed work, as reported in the preceding paper, has since been done on phytochrome effects in the natural environment, and it is encouraging to note that efforts are bring made to apply phytochrome research to horticulture (Decoteau, et al., 1993).
GREENHOUSES
As everyone interested in photoperiodism knows, L.H. Bailey (1891, 1892, 1893) used light from a Brush carbon-arc lamp to supplement natural light and extend the day in greenhouses. This was not; however, the first attempt to study the effects of electric light on plant growth. Carbon lamps* were used by Mangon (1861), and carbon arc lamps operated from steam or Otto gas engine driven Siemens, Grammes, or Alliance dynamos were used as early as 1853 and later by Siemens (1881), Deherain (1881), and Bonnier (1895). Prillieux (1869) investigated the effects of Drummond’s lamp** and gas light used for ordinary lighting on plant growth. Later Welsbach mantle incandescent gas light lamps*** (Corbett, 1899), neon (Hostermann 1922; Roodenburg, 1933), incandescent-filament lamps (Rane, 1894; Tjebbes and Uphof, 1921; Harrington, 1926; Truffaut and Thurneyssen, 1929), and quartz mercury lamps like the Cooper-Hewitt Uviarc were used for greenhouse supplementary light. Several early researchers noted the elongating effects of the greenhouse supplementary light, especially when incandescent-carbon or incandescent-filament lamps were used (Bonnier, 1895; Massart, 1920; Ramaley, 1931). Use of the term supplementary light (to supply what is lacking) is somewhat confusing, because in many cases the supplementary light was used continuously, throughout the night (Hostermann, 1922; Harrington, 1926), or only during the dark period (Cathey and Campbell, 1975), rather than as a supplement to natural light. In these studies, at least some of the growth effects reported are surely due to a response to the extremely long photoperiods, to end-of-day photomorphogenic effects, and to root zone warming rather than to additional photosynthesis.
* This was probably the Robert’s lamp introduced in 1852 in which a graphite rod was heated to incandescence in a vacuum or later in a nitrogen atmosphere.
** Drummond’s lamp, invented in 1826, heated a button of calcium oxide to incandescence. The resulting light was usually projected in a beam.
***Patented in 1886, Welsbach mantle lamps were made with a cotton wick impregnated with thorium oxide and a small amount of cerium oxide.
Using artificial light, usually from incandescent-filament lamps, for deliberate photoperiod control was initiated by Garner and Allard (1920) and was soon followed by many others. As photoperiod control became a production tool for floriculture and plant breeding, the more efficient fluorescent lamps were installed in a number of commercial greenhouses, often with unfortunate results; specifically failure or delay of flowering in long-day plants. Borthwick and Parker (1952) investigated this problem by comparing several kinds of fluorescent lamps, including special phosphor lamps, to incandescent lamps for efficiency in extending the greenhouse day to promote flowering of long-day plants. Annual beet and sugar beet flowered poorly or not at all under daylength extensions with any kind of fluorescent lamp, but flowered promptly when incandescent-filament light was used (Table 1). Although Odén, et al. (1932), Rasumov (1933), and Wenger (1934) had noted that the long wavelengths of light were necessary, or at least promotive, to normal flower stalk development, red radiation was considered the principal part of the spectrum controlling flowering. The action spectrum data probably influenced Borthwick and Parker (1952) to suggest that the much greater responsive-ness of plants to light from incandescent-filament than from fluorescent lamps was because the incandescent emitted a much greater percentage of red radiation than the fluorescent lamps. A few years later, of course, it was firmly established that the far-red emittion, or the lack thereof, had a strong influence on the response of plants to photoperiod control lighting.
TABLE 1. Effect of light source on flowering of beets. (Borthwick and Parker, 1955)
|
Photoperiod Control Light Source |
Annual Beets
Seed Stalks (per lot of 20) |
Sugar Beets
Flower Stalks (per lot of 12) |
| Incandescent
Fluorescent Warm White Soft White Cool White Daylight Agricultural* |
19
3 1 1 2 4 |
11
0 0 0 0 0 |
* Agricultural lamps emitted more red than the white lamps.
Many subsequent studies of photoperiodism compared daylength extensions obtained with fluorescent or incandescent light. Compared to fluorescent, the incandescent extension induced increased stem length in evergreen and deciduous tree species as well as herbaceous species such as tomato and soybean, promoted heading in millet, barley, and wheat, induced earlier flowering in Hyoscyamus niger, Petunia, dill, and other long-day species (Downs, et al., 1958; Downs and Hellmers, 1975; Vince-Prue, 1975), and produced greater pod set in H. niger (Table 2). Bulbing of onions was promoted by incandescent photoperiod control lighting and failed to occur when fluorescent was used (Woodbury and Ridley, 1969). Fluorescent photoperiod lighting failed to inhibit flowering of red-insensitive soybean varieties (Table 3), and when using photoperiod light to make a 13.5 h day for the most normal rate of reproduction in Ransom soybeans, incandescent lamps resulted in more pods than fluorescent lamps (Table 4).
TABLE 2. Reproduction of Hyoscyamus niger as affected by the source of light used to extend an 8-hour day in the greenhouse to 16 hrs.
| Photo-period
(h) |
Light Source | Duration
(d) |
Stem Length
(cm) |
Time to
Anthesis (d) |
Fruit Set
(%) |
| 8
16 16 |
none
Incandescent Fluorescent |
61
52* 61*
|
0.2
42 34 |
Vegetative
27 36 |
0
66 12 |
* Anthesis plus 25 days.
TABLE 3. Growth and reproduction of Blackhawk soybeans after 60 days under short days with various daylength extensions using incandescent or fluorescent light.
|
Light Regime |
Stem Length
(cm) |
Days to
Anthesis |
Pods > 2 cm in Length
Number Weight (mg) |
|
| 9 h
20 h Incandescent 20 h Fluorescent 20 h Incandescent and Fluorescent |
37
160 73
168 |
28
60 32
58 |
27
0 50
0 |
745
0 959
0 |
TABLE 4. Effect of the source of photoperiod control lighting on growth of Ransom soybeans in temperature-controlled greenhouses.
| Light Source | Stem Length
(cm) |
Leaf Area
(cm2) |
Fresh Weight
(g) |
Pod Number | Pod Weight
(g) |
| Incandescent
Fluorescent |
68
42 |
4859
2926 |
178.9
88.6 |
77
66 |
0.926
1.112 |
After the far-red reversibility of the red inhibition of hypocotyl growth in dark-grown seedlings was established (Downs, 1955), it was of interest to determine if this reversibility, and its confirmation of the activity of phytochrome, was also evident in internode growth of light-grown plants (Downs, et al., 1957). Irradiating bean plants for brief periods at the beginning of each dark period with far-red, so that the plants entered the dark period with phytochrome predominantly in the red-absorbing form, resulted in a large increase in internode length. The amount of elongation was proportional to the dark period remaining after the irradiation and was reversible by a subsequent exposure to red. Additional studies showed that what is now called ‘end-of-day’ far-red produced similar effects on most other bean varieties, sunflower, peanut, and morning glory. Also, end-of-day far-red promoted flowering of long-day plants, like dill, and short-day plants, such as millet (Downs, 1959) and milo (Lane, 1963), and had a marked effect on flowering of H. niger (Table 5). Extending the day with incandescent light is in effect providing end-of-day far-red, and the far-red effect becomes greater as the duration and irradiance of the incandescent light in increased.
TABLE 5. Effect of a long-day induction period with fluorescent light on promotion of flowering in Hyoscyamus niger by far-red at the close of 8-hour post-induction light periods.
| 10-day
Pretreatment Photoperiod |
Post-induction
Far-red (mins) |
Stem Length
(mm) |
Stage of
Flowering |
| 8 h
8 h 16 h 16 h |
0
5 0 5 |
0
0 13 43 |
0.0
0.0 3.0 6.0 |
High intensity discharge lamps are now widely used in greenhouses to supplement the low natural light levels of winter (Templing and Verbruggen, 1975; Duke, et al., 1975). Some researchers also use HID lamps, especially high pressure sodium (HPS) lamps, for photoperiod control lighting to prevent dormancy and to promote flowering of long-day plants. HPS lamps, however, are reported to be much less efficient than incandescent lamps, requiring a 4 to 8 fold increase in irradiance to provide the same photoperiodic stimulus as the 1:1 red/far-red ratio* of incandescent lamps (Cathey and Campbell, 1964). In fact, the benefits of HPS supplemental light is enhanced by the addition of some incandescent lamps (Cathey and Campbell, 1977).
Today the incandescent lamp remains the chief source of light for photoperiod control because it is well established that a red/far-red ratio of 0.671** is more effective than the 7.6969 ratio from fluorescent or the 2.7 of HPS and 2.5 ratio of MH high intensity discharge lamps.
* The red/far-red ratio of incandescent lamps is more nearly 0.67 than 1
** 640-660/720-740 nm
PLANT GROWTH CHAMBERS
Early attempts to use electric lamps as the sole source of light for plant growth chiefly used nitrogen-filled incandescent-filament lamps, the Mazda C lamp (Harvey, 1922; Maximov, 1925; Davis and Hoagland, 1928; Sande-Bakhuyzen, 1928; Redington, 1929; Truffaut and Thurneyssen, 1929; Stoughton, 1930; Arthur et al, 1930; Steinberg, 1931; Bracket and Johnston 1932; Johnston, 1932; Wilson, 1937; Wettstein and Pirschle, 1940), although some of these efforts utilized neon, low-pressure sodium, mercury tungsten, mercury arc, mercury vapor, or carbon-arc light, alone or in conjunction with incandescent lamps in order to increase the illuminance (Roodenburg, 1931; Johnston, 1938; Steward and Arthur, 1934; Weigel and Knoll, 1936; Pirschle and Wettstein, 1940; Ullrich, 1941; Aberg, 1941, 1943). Several of these examples where incandescent light was used noted excessive stem elongation. Roodenburg (1940) stated that near infra red produces a specific elongation effect and Aberg (1943) in noting the elongation that occurred, concluded that “The infra-red rays of shorter wavelength that penetrate a layer of water 3 cm in thickness probably have a favorable effect on the internode elongation in the tomato plant.”
In addition to the etiolation, a major problem with these early efforts was the low light level, equivalent to about 160 μmols m-2 s-1 and often less, generated by these lamps. In order to obtain a higher illuminance, Mitchell (1935) installed a new type, high intensity, carbon-arc lamp for respiration and photosynthesis studies. These lamps had been designed for use in hospital solaria to treat extrapulmonary tuberculosis patients. Gains made by the patients during summer exposure to sunlight were lost during winter months due to low light levels and cloudy days. Clinical sunlight, recommended at 140-160 mW m-2 between 290 and 310 nm, could be and was supplied by these carbon arc lamps (Grieder and Downes, 1932).
E.J. Kraus and Jack Mitchell left the University of Chicago about 1935 to join the Beltsville photoperiod project. Thus it was probably at their recommendation that the four temperature- and humidity-controlled plant growth chambers that were installed at Beltsville about 1937 were equipped with these carbon-arc lamps . Soybeans grown under the arc lamps consistently had a lower carbohydrate content than plants grown in the greenhouse. Parker and Borthwick (1949) concluded that the low carbohydrate level probably resulted from the small amount of red radiation emitted by the ‘Sunshine’ carbons in these carbon-arc lamps. So the following year they installed incandescent lamps that provided 8 to 10% of the illuminance of the arc lamp to provide additional red radiation. Soybeans grown under carbon-arc light plus incandescent revealed an increase in starch and sugars that could not be accounted for by the small increase in illuminance (Table 6). In retrospect it seems strange that Parker and Borthwick (1949) would attribute these gains to the increase in red due to the incandescent since a much larger increase in red obtained by using a different type carbon, .025 carbons, had very little effect. (Table 6).
TABLE 6. Carbohydrate composition of Biloxi soybeans grown for 4 weeks under a carbon-arc lamp utilizing different carbon types, with and without incandescent lamps. (Parker and Borthwick, 1949)
| Carbon Type | Red (%)* | Reducing Sugars | Sucrose | Starch |
| (mg per plant) | ||||
| Sunshine Carbons | 42 | 32.5 | 6.6 | 39 |
| Sunshine Carbons + Incand | 49 | 73.0 | 14.0 | 70 |
| Sunshine Carbons + 025 Carbons | 51 | 45 | 8.0 | 36 |
* 650 nm as percent of 450 nm radiation.
These carbon arc/incandescent lighted chambers were kept in almost continuous use for over 30 years, but operational and maintenance problems induced Parker and Borthwick (1950) in 1947 to begin planning a controlled-environment room lighted with fluorescent lamps. Several years earlier fluorescent lamps had been tested satisfactorily for plant growth (Naylor and Gerner, 1940; Hartmann and McKinnon, 1943; Hamner, 1944; Went, 1944), but the low illuminance available from these lamps was inadequate for controlled-environment rooms. The introduction of the 8 ft. slimline lamp following World War II seemed to provide a means of obtaining sufficient illumination for plant growth over relatively large areas, especially when the lamp current was increased from 200 to 300 mA. During the design phase of this room, Parker compared plant growth under slimline fluorescent with and without incandescent supplementary light. As with the carbon-arc lamp rooms, the avowed purpose of the incandescent lamps was to increase a possible deficiency of red radiation. The supplementary incandescent light resulted in an 18% increase in dry weight. Withrow and Withrow (1947) had reported that adding incandescent to fluorescent light increased yield, and later reports verified the increased growth due to added incandescent light (Dunn and Went, 1959; Helson, 1965; Deutch and Rasmussen, 1974; Cathey et al, 1978). Dunn and Went (1959) noted that the effect of adding 10% of the fluorescent illuminance with incandescent was no greater when added to red fluorescent than to blue fluorescent, concluding that “while the most obvious explanation is that the effect is due to the infra red radiation of the incandescent lamps, it is unlikely that the far-red and infra red rays of the incandescent light was responsible (for the increased growth) since they would have been more effective when added to blue than to the red fluorescent light”.
Parker planned additional experiments to evaluate the plant growth effectiveness of various kinds of fluorescent lamps, including experimental lamps with special phosphors like the Agricultural, and to examine other quantities of incandescent supplementary light. The results of these studies were never published, but when the fluorescent-lighted room was completed about 1950 it contained cool white fluorescent and incandescent lamps that provided about 10% of the illuminance of the fluorescent lamps. In order to facilitate future plant growth chamber construction, Joe Ditchman, a GE engineer assigned to biological lighting development, calculated that 10% of the illuminance of the slimline fluorescent lamps could be obtained by installing incandescent lamps at the rate of 30% of the fluorescent wattage. Due to lack of data on plant response to other levels of incandescent supplementary light, this value, 30% of the installed fluorescent watts, became a guideline for use in growth chamber design. The validity of this percentage, of course, was lost as designers increased the efficiency of the fluorescent lamp. For example about 1963, a chamber was constructed at Beltsville using 1500 mA, non-circular cross section, fluorescent lamps and, while the incandescent effect was still apparent at light levels as high as 500 μmols m-2 s-1 (Table 7), increasing the intensity of the main light source decreases the incandescent effect of the ‘standard’ incandescent installation. This fact was also noted by Meijer (1957) and Sanchez and Cogliatti (1975). Thus it is not surprising to find that increasing the percentage of incandescent watts increases the incandescent effect in chambers lighted with 1500 mA lamps (Krizek and Ormrod, 1980; Murakami, et al., 1991).
TABLE 7. Effect of radiation from incandescent lamps during the fluorescent light period on growth of Ransom soybeans after 32 days
| Main Axis | Branches | |||
| Light Source | Length
(cm) |
Leaf Area
(cm2) |
Number | Leaf Area
(cm2) |
| Fluorescent
Fluorescent plus Incandescent |
77
93 |
1235
1410 |
8
5 |
1993
1622 |
In addition to increased plant weight (Rajan, et al., 1971; Deutch and Rasmussen, 1974; Hurd, 1974), the addition of incandescent light to the fluorescent system also resulted in increased flower weight and number of florets in Chrysanthemum, while reducing the number of days required to develop flower color (Hassan and Newton, 1975) and improved flowering of long-day plants (De Lint, 1958; Friend et al 1961; Dietzer et al, 1979). The incandescent light may also increase stem elongation, alter leaf area, and reduce branch and tiller development (Rajan, et al., 1971; Summerfield and Huxley, 1972; Proctor, 1973; Deutch and Rasmussen, 1974; Downs and Thomas, 1990; Casal, et al., 1985). Moreover, if the incandescent and fluorescent lamps are not turned off simultaneously a substantial, often undesirable, stem lengthening can occur (Table 8) that may not be recognized by many plant growth chamber users as an end-of-day far-red effect. With some plants incandescent light is essential for normal plant development (Friend, et al., 1961), but it is also clear that with other plants incandescent light is a major factor in the inability to simulate the field phenotype (Tanner and Hume, 1976).
TABLE 8. Effect of light quality for a 30 minute period after the close of the high-intensity light period on growth of tobacco seedlings.
| Variety | Light source | Stem | Fifth Leaf | |
| Length | Length | Width | ||
| (cm) | (cm) | (cm) | ||
| Coker 319 | Fluorescent | 6.3 | 9.7 | 16.2 |
| Incandescent | 13.7 | 10.0 | 19.7 | |
| NC2326 | Fluorescent | 5.7 | 9.0 | 15.8 |
| Incandescent | 10.0 | 10.2 | 19.5 | |
While fluorescent-lighted chambers have been constructed without incandescent supplemental light (Doorenbos, 1964), the advantages of using the incandescent to increase growth, accelerate flowering in long-day plants, control flowering in red-insensitive varieties, and produce end-of-day far-red effects makes their addition in fluorescent-lighted plant growth chambers extremely useful and in some cases indispensable. For example tissue cultures of Loblolly pine fail to differentiate without incandescent light added to the fluorescent system.
In other cases where the incandescent supplemental light is a detriment to obtaining the growth or plant habit desired, the problem can be solved, in soybeans at least, by utilizing correct photoperiod regimes and/or using the incandescent lamps correctly (Downs and Thomas, 1990). In other examples of inadequate plant development, the incandescent lamps can be easily turned off.
HIGH INTENSITY DISCHARGE LAMPS
High intensity discharge lamps in the form of mercury or phosphor-coated mercury (sometimes called mercury-fluorescent) lamps were added to the fluorescent-incandescent system as early as 1955 (Oda, 1962). The development of similar systems by others soon followed (Leiser, et al., 1960; Yamamoto, 1970); each apparently without knowledge of the other installations. Chambers lighted solely with phosphor-coated mercury lamps also were constructed (Bretschneider-Hermann, 1964; Chandler, 1972; Smeets, 1978), but the low efficiency of these lamps limited their use. When the highly efficient metal halide lamps were introduced, plant growth chamber designers quickly incorporated them into new chambers (Nakamura, 1972; Kawarda and Shibata, 1972; Warrington, et al., 1976; Eguchi, 1986) and ultimately retrofitted them into older chambers (Downs, 1988). The further increase in light-producing efficiency achieved by the introduction of the high pressure sodium lamp about 1965 resulted in a number of trials with this light source (Downs and Hellmers, 1975). In our studies, HPS proved less than satisfactory as a sole source of light for field crop plants, but plants grew well when irradiated with a 1:1 mixture of mercury, or metal halide and HPS. In contrast to our earlier results, Smeets at Wageningen designed a 100 m2 room with only HPS lamps that appears to provide satisfactory growth of several floricultural crops (personal observation).
Although HID lamps can provide the same irradiance as fluorescent lamps at a substantially reduced power requirement, the chief reason for using them seems to be to increase the PPFD above that normally available from fluorescent lamps. An exception is the work at the Climate Lab in New Zealand, which was primarily interested in obtaining a spectral distribution equivalent to sunlight including an appropriate red/far-red ratio (Warrington, et al., 1976; Warrington, et al., 1978), and was only secondarily interested in super high light levels. Today we see chambers being constructed with light levels equalling or exceeding peak solar radiation. The reason given for the high irradiance is usually that it is necessary for simulating field studies. It would seem that the R/FR ratio of natural light would also be a requirement for simulating field studies, but this subject is rarely encountered in arguments for artificially-produced solar irradiance levels. The spectral distributions of the tin chloride lamp, which was never produced commercially, and the Tungsram daylight metal halide containing dysporsium (Tischner and Vida, 1981) come very close to matching the natural light spectral distribution.
A question that arises frequently in the design of HID-lighted growth chambers is whether incandescent lamps should be added. Tibbitts, et al., (1987) reported that incandescent lamps had little to no effect on growth of mustard and wheat when they were added to high intensity discharge lamps. However, there was a small but significant increase in soybean vegetative growth (Fig, 1), and Casal, et al. (1985) reported that incandescent light reduced tillering and advanced reproductive development in Lolium.
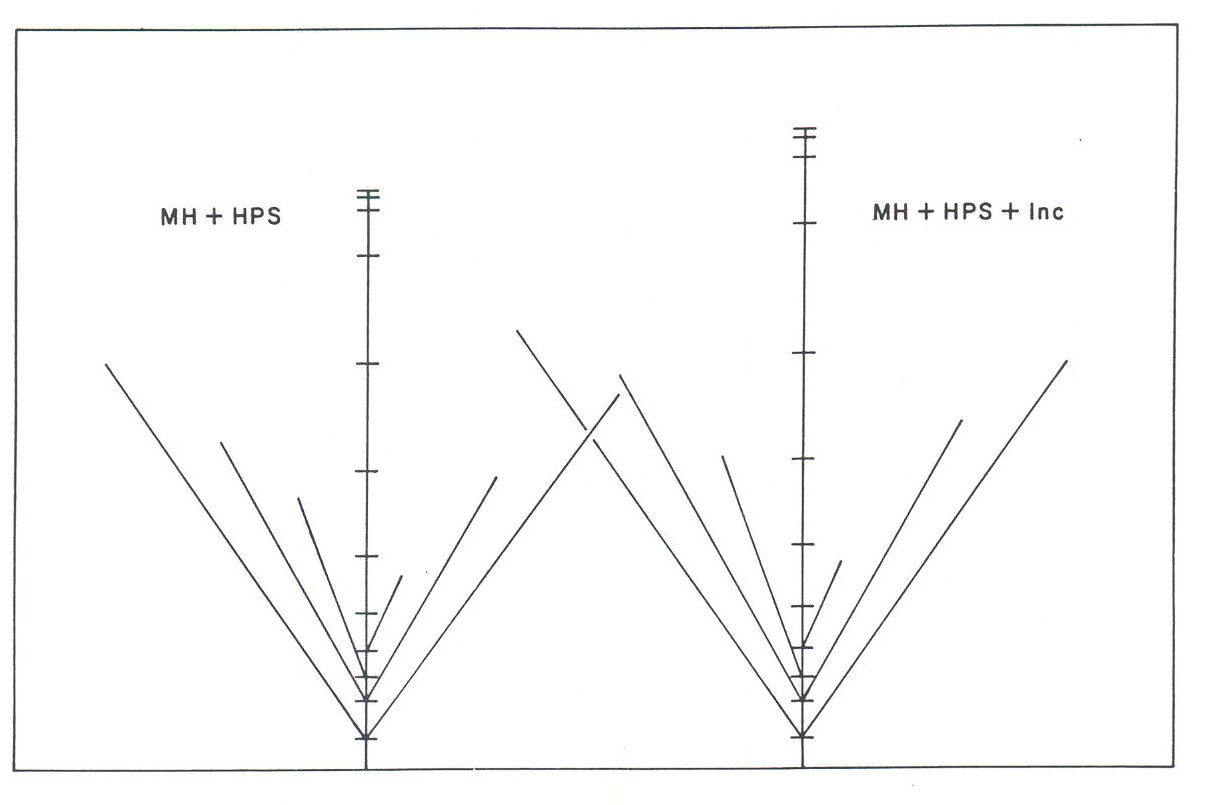
Fig. 1. Schematic of Ransom soybean growth after 30 days under a 1:1 ratio of high pressure sodium and metal halide lamps with and without incandescent.
We originally assumed that the lack of far-red effect when incandescent lamps were added to HID-lighted chambers was due to the higher HID irradiance. This is not a satisfactory explanation, however, since a marked far-red effect failed to be discernable at HID light levels comparable to fluorescent-lighted rooms (less than 500 μmols m-2 s-1). Part of the problem seems to be that incandescent lamps provide red as well as far-red;and thus, the net increase in far-red relative to red is not as great as might be assumed. For example, the red/far-red ratio in a reach-in chamber lighted with 16, 115-W VHO fluorescent lamps was 6.684. Adding incandescent at an input wattage of 33% of the installed fluorescent watts reduced the R/FR ratio to 1.884. When we retrofitted this chamber with HID lamps the R/FR ratio was 2.526 with MH and 2.749 with a 1:1 mixture of MH and HPS lamps. Adding incandescent decreased the ratio to 1.7 and 2, respectively. These ratios are similar to those from fluorescent-incandescent systems but the far-red effect is much less.
In part, this lack of an incandescent effect can be alleviated by increasing the incandescent lamp wattage (Warrington, 1978) to equal that of the HID lamps. While a properly designed reflector and ventilation system can remove the thermal radiation from the HID lamps (Downs, 1989), the large amount of long wavelength radiation resulting from such a large wattage of incandescent lamps makes a water filter essential. Unfortunately, the water filter is often not practical because it increases design costs and requires much more maintenance than the typical lamp loft barrier. The heat removal problem might be avoided by adding far-red without any increase in red radiation. In theory this could be done by using blue incandescent lamps which have a red/far-red ratio of 0.004 compared to the 0.671 of white incandescent ones, but in practice the far-red effect from blue incandescent lamps added to HID lamps is about the same as with white incandescent lamps.
Plants grown under HID lamps often produce abnormally short internodes, a fact observed by Warrington et al (1978), even when incandescent lamps were added. End-of-day exposures to incandescent lamps can be used as a tool to increase internode lengths to more acceptable values (Table 9). End-of-day irradiations with blue incandescent lamps, however, produce excessive elongation. (Table 10). Also, using the incandescent lamp for dark period interruptions, as an end-of-day treatment, or for daylength extensions can accelerate flowering of many long-day plants and control flowering of red-insensitive soybeans. The evidence seems to favor the addition of incandescent lamps to HID systems.
TABLE 9. Oregon 91 snapbeans grown under MH and HPS lamps with and without 30 min end-of-day incandescent irradiation.
|
Light Source |
Stem Length
(cm) |
Branch Length
(cm) |
Leaf Area
(cm-2) |
Top Weight
(g) |
| Incandescent
No Incandescent |
31.0
11.4 |
33
19 |
814
626 |
32.86
25.78 |
TABLE 10. Seneca chief squash grown under MH and HPS lamps with 15 min. end-of-day exposures to white or blue incandescent lamps.
| Length | Length | ||||
|
Light Source |
Hypocotyl
(cm) |
Stem
(cm) |
Petiole
1st Leaf |
Leaf Area
(cm2) |
Top Weight
(g) |
| White Incandescent
Blue Incandescent |
1.6
5.3 |
2.7
7.6 |
12.8
29.4 |
514
388 |
26.53
32.77 |
And thus, it is recommended that the design and construction of plant growth chambers continue to contain a provision for utilization of the incandescent lamp as part of the total irradiance system, to be implemented at the discretion of the investigator to meet the phytochrome requirements of the various biological organisms that may be grown in the chamber.
REFERENCES
Aberg, B. 1943. Physiologische und ökologische Studien über dir Pflanzliche Photomorphose. Symp. Bot. Upsaliensis 8:1-189.
Arthur, J.M., J.D. Guthrie, and J.M. Newell. 1930. Some effects of artificial climates on the growth and chemical composition of plants. Am. J. Bot. 17:416-482.
Bailey, L.H. 1891. Some preliminary studies on the influence of the electric light upon greenhouse plants. Cornell Univ. Agri. Expt. Sta. Bull. 30:83-122.
Bailey, L.H. 1892. Second report on electroculture. Cornell Univ. Agri. Expt. Sta. Bull. 42:199-212.
Bailey, L.H. 1893. Third report on electroculture. Cornell Univ. Agri. Expt. Sta. Bull. 55:147-157.
Bonnier, G. 1895. Influence de la lumière électrique continue sur la forme et la structure des plantes. Rev. Gén. Bot. 7:241-257; 289-306; 332-342; 409-419.
Borthwick, H.A. and M.W. Parker. 1952. Light in relation to flowering and vegetative development. Rept. 13th. Internatl. Hort. Congress, London.
Bracket, F.S. and E.S. Johnston. 1932. The function of radiation in the physiology of plants. I. General methods and apparatus. Smithsonian Misc. Coll. 87:1-10.
Bretschneider-Herrmann, R. 1964. Phytotron in Rauisch-Holzhausen. Technical details and experiences. p. 24-26. In: P. Chouard and N. de Bilderling (eds). Phytotronique I. Centre Natl. Recherche Sci. Paris.
Casal, J.J., V.A. Deregibus, and R.A. Sanchez. 1985. Variations in tiller dynamics and morphology in Lolium multiflorum Lam. vegetative and reproductive plants as affected by differences in red/far-red irradiation. Ann. Bot. 56:553-559.
Cathey, H.M. and L.E. Campbell. 1964. Lamps and lighting: a horticultural review. Lighting Design Applic. Nov:1-12.
Cathey, H.M. and L.E. Campbell. 1975. Effectiveness of five vision-lighting sources on photo-regulation of 22 species of ornamental plants. J. Am. Soc. Hort. Sci. 100:65-71.
Cathey, H.M. and L.E. Campbell. 1977. Plant productivity: New approach to efficient light sources and environmental control. Trans. Am. Soc. Agri. Eng. 20:260-266.
Cathey, H.M., L.E. Campbell, and R.W. Thimijan. 1978. Comparative development of 11 plants grown under various fluorescent lamps and different durations of irradiation with and without additional incandescent lighting. J. Am. Soc. Hort. Sci. 103:781-791.
Chandler, B. 1972. Towards a simple growth room design. p 279-288. In: P. Chouard and N. de Bilderling (eds). Phytotronique et Prospective Horticole. Gauthiers-Villars, Paris.
Corbett, L.C. 1899. A study of the effect of incandescent gas light on plant growth. West Va. Agri. Expt. Sta. Bull. 62:77-110.
Davis, A.R. and D.R. Hoagland. 1928. An apparatus for the growth of plants in a controlled environment. Plant Physiol. 3:277-292.
Decoteau, D.R, H.A. Hatt, J.W. Kelly, M.J. McMahon, N. Rajapakse, R.E. Young, and R.K. Pollack. 1993.Applications of photo-morphogenesis research to horticultural systems. HortScience 28:974, 1063.
Deherain, P.P. 1881. Expériences sur l’influence qu’exerce la lumière électrique sur la dévellopement des végétaux. Ann. Agron. Paris 7:551-575.
Deitzer, G., R. Hayes, and M. Jabben. 1979. Kinetics and time dependence of the far-red light on the photoperiodic induction of flowering in Wintex barley. Plant Physiol. 64:1015-1021.
DeLint, P.J.A.L. 1958. Stem formation in Hyoscyamus niger under short days including supplementary irradiation with near infra red. Meded. Lanbouwhogesch. Wageningen. 58:1-5.
Deutch, B. and O. Rasmussen. 1974. Growth chamber illumination and photomorphogenic efficacy. I. Physiological action of infra red radiation beyond 750 nm. Physiol. Plant. 30:64-71.
Doorenbos, J. 1964. The phytotron of the Laboratory of Horticulture, State Agricultural College, Wageningen. Meded. Dir. Tuinbou. 27:432-437.
Downs, R.J. 1955. Photoreversibility of leaf and hypocotyl elongation of dark-grown Red Kidney bean seedlings. Plant Physiol. 30:468-473.
Downs, R.J. 1959. Photocontrol of vegetative growth. p. 129-135. In: R.B. Withrow (ed). Photoperiodism and Related Phenomena in Plants and Animals. Am. Assoc. Adv. Sci. Washington, D.C.
Downs, R.J. 1988. Retrofitting plant growth chambers with high intensity discharge lamps. Paper 88-4016. Am. Soc. Agri. Eng. St. Josephs, MI.
Downs, R.J. 1989. Reflector design for HID lamps used in plant growth chambers. Paper 89-4581. Am. Soc. Agri. Eng. St. Josephs, MI.
Downs, R.J. and H. Hellmers. 1975. Environment and the Experimental Control of Plant Growth. Academic Press, London.
Downs, R.J. and J.F. Thomas. 1990. Morphological and reproductive development of soybeans under artificial conditions. Biotronics 19:19-32.
Downs, R.J., H.A. Borthwick, and S.B. Hendricks. 1957. Photoreversible control of elongation of Pinto beans and other plants under normal conditions of growth. Bot. Gaz. 118:119-208.
Downs, R.J., H.A. Borthwick, and A.A. Piringer. 1958. Comparison of incandescent and fluorescent lamps for lengthening photoperiods.Proc. Am. Soc. Hort. Sci. 71:568-578.
Duke, W.B., R.D. Hagi, J.F. Hunt, and D.L. Linscott. 1975. Metal halide lamps for supplemental lighting in greenhouses: crop responses and spectral distribution. Agron. J. 67:49-53.
Dunn, S. and F.W. Went. 1959. Influence of fluorescent light quality on growth and photosynthesis of tomato. Lloydia 22:302-324.
Eguchi, H. 1986. Biotron Institute, Kyushu University. Japan.
Friend, D.J., V.A. Helson, and J.E. Fisher. 1961. The influence of the ratio of incandescent and fluorescent light on flowering response of Marquis wheat grown under controlled condition. Canad. J. Plant Sci. 41:418-427.
Garner, W.W. and H.A. Allard. 1920. Effect of the relative length of day and night and other factors of the environment on growth and reproduction of plants. J. Agri. Res. 18:553-606.
Grieder, C.E. and A.C. Downes. 1932. The carbon arc as a source of sunshine, ultraviolet, and other radiation. Trans. Illum. Eng. Soc. 27:637-653.
Hamner, K.C. 1944. Description of a chamber for growing plants under controlled conditions. Bot. Gaz. 105:437-441.
Harrington, J.B. 1926. Growing wheat and barley hybrids in winter by means of artificial light. Sci. Agri. 7:125-130.
Hartmann, H.T. and L.R. McKinnon. 1943. Environment control cabinets for studying the inter-relation of temperature and photoperiod on the growth and development of plants. Proc. Am. Soc. Hort. Sci. 42:475-480.
Harvey, R.B. 1922. Growth of plants in artificial light from seed to seed. Science 56:366-367.
Hassan, M.R.A. and P. Newton. 1975. Growth of Chrysanthemum morifolium cultivar Pollyanna in natural and artificial light. p. 154-164. In: P. Chouard and N. de Bilderling (eds). Phytotronics in Agricultural and Horticultural Research. Gauthier-Villars, Paris.
Helson, V.A. 1965. Comparison of GroLux and cool white fluorescent lamps with and without incandescent as light sources used in plant growth rooms for growth and development of tomato plants. Canad. J. Plant Sci. 45:461-466.
Hostermann, G. 1922. Kulturversuche mit elektrischen Licht. Gartenwelt. 26:74-75.
Hurd, R.G. 1974. The effect of incandescent supplement on the growth of tomato plants in low light. Ann. Bot. 38:613-123.
Johnston, E.S. 1932. The function of radiation in the physiology of plants. II. Some effects of near infra red radiation on plants. Smithsonian Misc. Coll. 87:1-16.
Johnston, E.S. 1938. Plant growth in relation to wavelength balance. Smithsonian Misc. Coll. 97:1-18.
Kawarda, A. and K. Shibata. 1972. Phytotron, Institute of Physical and Chemical Research. p. 87-91. In: A. Funada et al (eds) Phytotrons and Growth-Cabinets in Japan. Japanese Society Environ. Control in Biol.
Krizek, D.T. and D.P. Ormrod. 1980. Growth responses of ‘Grand Rapids’ lettuce and ‘First Lady’ Marigold to increased far-red and infrared radiation under controlled conditions. J. Am. Soc. Hort. Sci. 105:936-939.
Lane, H.C. 1963. Effect of light quality on maturity in the milo group of Sorghum. Crop Sci. 3:496-499.
Leiser, A.T., A.C. Leopold, and A.L. Shelly. 1960. Evaluation of light sources for plant growth. Plant Physiol. 35:392-395.
Mangon, Hervé M. 1861. Production de la matière verte des feuilles sous l’influence de la lumière électrique. Compt Rend. Acad. Sci. 53:243-244.
Massart, J. 1920. L’action de la lumière continue sur la structure des feuilles. Acad. Roy. Belg. Bull. Cl. Sci. 6:37-43.
Maximov, N.A. 1925. Pflanzenkultur bei elektrischen Licht und ihre Anwendung bei Samenprüfung und Pflanzenzüchtung. Biol. Zentralbl. 45:627-639.
Meijer, H. 1957. The influence of light quality on the photoperiodic response of Salvia occidentalis. Acta Bot. Neerl. 7:801-805.
Meijer, G. 1971. Some aspects of plant irradiation. Acta Hort. 22:102-108.
Mitchell, J.W. 1935. A method of measuring respiration and carbon fixation of plants under controlled environmental conditions. Bot. Gaz. 97:376-387.
Murakami, K., K. Horiguchi, M. Morita, and I. Aiga. 1991. Growth control of sunflower (Helianthus annuus L. cv Russian Mammoth) seedlings by additional far-red radiation. Environ. Control in Biol. 29:73-79.
Nakamura, M. 1972. Environment-controlled room, Hatano Tobacco Experiment Station. p. 81-82. In: S. Funada, et al (eds). Phytotrons and Growth Chambers in Japan. Japanese Soc. Environ. Control in Biol.
Naylor, A.W. and G. Gerner. 1940. Fluorescent lamps as a source of light for growing plants. Bot. Gaz. 101:715-716.
Oda, Y. 1962. The air-conditioned darkroom with artificial light. p. 18-21. In: S. Matsumura (ed). Environment-controlled Growth Rooms in Japan. Fac. Agri. University Tokyo.
Odén, S., G. Köhler, and G. Nilsson. 1932. Plant cultivation with the aid of electric light. Proc. Internatl. Illum. Cong. 2:1298-1326.
Parker, M.W. and H.A. Borthwick. 1949. Growth and composition of Biloxi soybean grown in a controlled environment with radiation from different carbon-arc sources. Plant Physiol. 24:345-358.
Parker, M.W. and H.A. Borthwick. 1950. A modified circuit for slimline fluorescent lamps for plant growth chambers. Plant Physiol. 25:86-91.
Parker, M.W., S.B. Hendricks, and H.A. Borthwick. 1950. Action spectrum for the photoperiodic control of flower initiation of the long day plant Hyoscyamus niger. Bot. Gaz. 111:242-252.
Pirschle, F. and K. von Wettstein. 1940. Einige vorläufige Beobachtungen über die Wirkung verscheidener Lichtintensitäten und -qualitäten auf höhere Pflanzen unter konstanten Bedingungen. Biol. Zentralbl. 60:626-658.
Prillieux, E. 1869. De l’influence de la lumière artificielle sur la rèduction de l’acide carbonique par les plantes. Compt. Rend. Acad. Sci. 68:408-412.
Proctor, J.R.A. 1973. Developmental changes in radish caused by brief end of day exposures to far-red radiation. Canad. J. Bot. 51:1075-1077.
Rajan, A.K., B. Betteridge, and G.E. Blackman 1971. Interrelationships between the nature of the light source, ambient air temperature and the vegetative growth of different species within growth cabinets. Ann. Bot. 35:32:323-342.
Ramaley, F. 1931. Growth of plants under continuous light. Science 73:566-567.
Rane, F.W. 1894. Electroculture with the incandescent lamp. West Va. Agri. Expt. Sta. Bull. 37:1-27.
Rasumov, V.I. 1933. The significance of the quality of light in photoperiodical response. Bull. Appl. Bot. Genet. Plant Breed. 3:217-251.
Redington, G. 1929. A study of the effect of diurnal periodicity upon plant growth. Trans. Roy. Soc. Edinburgh 56:247-272.
Roodenburg, J.W.M. 1931. Künstlichkultur. Angew. Bot. 13:162-166.
Roodenburg, J.W.M. 1933. Pflanzenbestrahlung mit Neonlicht. Schwizer Elektro Rundschau VII.
Roodenburg, J.W.M. 1940. Das Verhalten von Pflanzen in verschiedenen-farbigen Licht. Rec. Trav. Bot. Neerl. 37:301-376.
Sanchez, R.A. and D. Cogliatti. 1975. The interaction between phytochrome and white light irradiance in the control of leaf shape in Taraxacum officinale. Bot. Gaz. 136:281-285.
Sande-Bakhuyzen, H.L. van de. 1928. Studies of wheat grown under constant conditions. Plant Physiol. 3:1-6; 7-30.
Siemens, M.C.W. 1881. On the influence of light upon vegetation and on certain physical principles involved. Proc. Roy. Acad. 30:210-219 and 293.
Smeets, L. 1978. IVT Phytotron, 1953-1978. Neth. J. Agri. Sci. 26:1-132.
Steinberg, R.A. 1931. An apparatus for growing plants under controlled environment conditions. J. Agri. Res. 43:1071-1084.
Steward, W.D. and J.M. Arthur. 1934. Some effects of radiation from a quartz mercury vapor lamp upon mineral composition of plants. Contr. Boyce Thompson Inst. 6:225-245.
Stoughton, R.H. 1930. Apparatus for the growing of plants in a controlled environment. Ann. Appl. Biol. 17:90-106.
Summerfield, R.J. and P.A. Huxley. 1972. Management and plant husbandry problems of growing soyabean and cowpea cultivars under artificial light. Rept. Internatl. Inst. Tropical Agri. Tropical Grain Legume Physiol. Proj.
Tanner, J.W. and D.J. Hume. 1976. The use of growth chambers in soybean research. p. 342-351. In: L.D. Hill (ed). World Soybean Research. Interstate Publ. Danville, Va.
Templing, B.C. and M.A. Verbruggen. 1975. Lighting Technology in Horticulture. Philips Gloeilampenfabrieken, Eindoven, Neth.
Tibbitts, T.W., D.C. Morgan, and I.J. Warrington. 1987. Growth of lettuce, spinach, mustard, and wheat plants under four combinations of high pressure sodium, metal halide and tungsten halogen lamps at equal PPFD. J. Am. Soc. Hort. Sci. 108:622-630.
Tischner, T.W. and D. Vida. 1981. Metal halide lamps with rare earth additives for plant growth tests. Tungsram Tech. Rev. 48:1889-1895.
Tjebbes, K. and J.C. Uphof. 1921. Der Einfluss des elektrischen Lichtes auf das Pflanzenwachstum. Landwirt. Jahrb. 56:315-328.
Truffaut, G. and G. Thurneyssen. 1929. Influence de la lumière artificielle sur la croissance des plantes supèriéures. Compt. Rend. Acad. Sci.
Ullrich, H. 1941. Zur Frage der Entwicklung der Pflanzen bei ausschliesslich künstlicher Beleuchtung. Ber. Bot. Ges. 59:192-232.
Vince-Prue, D. 1975. Photoperiodism in Plants. McGraw-Hill, London.
Warrington, I.J. 1978. Controlled environment lighting – high pressure discharge lamps-based systems. Proc. Growth Chamber Environ. Symp. 20th Internatl. Hort. Congress, Sydney, Australia.
Warrington, I.J., K.J. Mitchell, and C. Halligan. 1976. Comparison of plant growth under four different lamp combinations and various temperature and irradiance levels. Agric. Meteorol. 16:231-245.
Warrington, I.J., E.A. Edge, and L.M. Green. 1978. Plant growth under high radiant energy fluxes. Ann. Bot. 42:1305-1313.
Weigel, R.G. and O.H. Knoll. 1936. Lichtbiologische Beeinflussung der Aufzucht von Gemüsepflanzen. Das Licht 6:219-261.
Wenger, R. 1934. Some effects of supplementary illumination with Mazda lamps on the carbohydrate and nitrogen metabolism of the Aster. p. 11. Abstr. 11th Ann. Meeting Plant Physiol. Soc.
Went, F.W. 1944. Plant growth under controlled conditions. II. Am. J. Bot. 31:135-150.
Wettstein, F. von and K. Pirschle. 1940. Klimakammern bei konstanten Bedingungen für die Kultur höhere Pflanzen. Naturwissenschaften 28:537-543.
Wilson, A.R. 1937. An apparatus for growing plants under controlled environmental conditions. Ann. Appl. Biol. 24:911-931.
Withrow, R.B. and A.P. Withrow. 1947. Plant growth with artificial sources of radiant energy. Plant Physiol. 22:494-513.
Woodbury, G.W. and J.R. Ridley. 1969. The influence of incandescent and fluorescent light on bulbing response of three onion varieties. J. Am. Soc. Hort. Sci. 94:365-367.
Yamamoto, T. 1970. The Hokkaido National Agricultural Experiment Station Phytotron. Japan Res. Quarterly 5:52-58.
Downs, R.J. 1994. History and applications in controlled environments, p 69-87. In: T.W.Tibbitts (ed.). International Lighting in Controlled Environments Workshop, NASA-CP-95-3309.
Copyright © March 1994 NASA [National Aeronautics and Space Administration].
All rights reserved.